Q-omics provides the consensus-scored SVIP profile across patient tissues and cancer cell-line models. SVIP expression is associated with patient survival in 23 of 34 cancer types, with the highest sampling consensus in HNSC. Among the 18 cancer types available for tumor–normal comparison, SVIP is differentially expressed in 13, with the highest sampling consensus in KIRC. Additionally, SVIP protein abundance shows 23,007 significant protein co-abundance associations, with the highest sampling consensus in GBM. Together, these results highlight HNSC, KIRC, and GBM as cancer lineages where SVIP shows reproducible signals across survival, tumor–normal expression, and patient cross-omics analyses.
Every result is evaluated using two consensus scores. Sampling consensus measures how consistently a finding is reproduced within a cancer lineage across different conditions. Lineage consensus measures how broadly the result is shared across cancer types, distinguishing pan-cancer signals from lineage-specific patterns.
Premium analyses for SVIP — synthetic lethality, tumor antigen, and pembrolizumab response.
This table summarizes SVIP survival associations across molecular data types. SVIP RNA expression shows survival associations in the most cancer types (23), followed by mutation status (1) and mass-spec protein abundance (4). The rightmost column indicates the cancer type with the highest sampling consensus for each molecular layer.
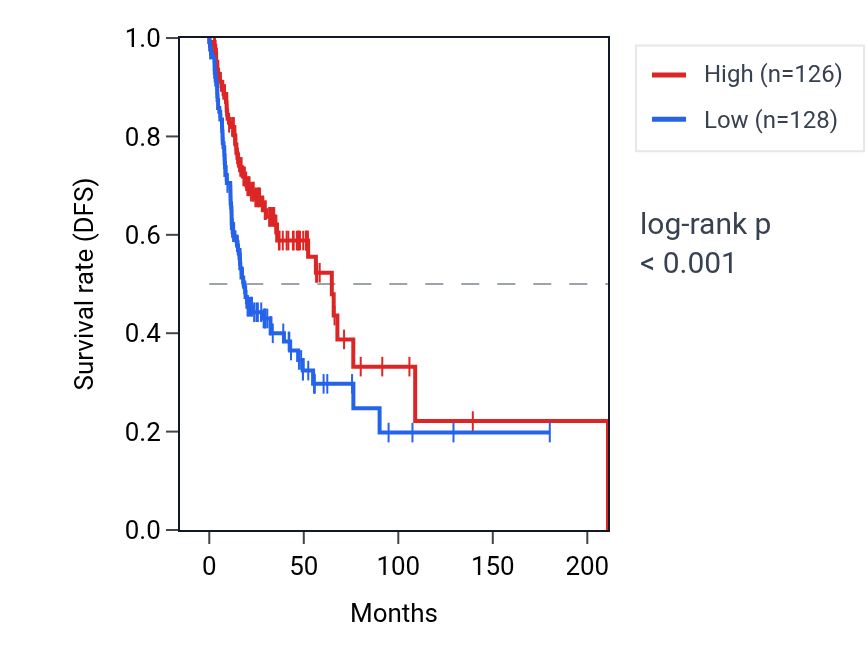
This table ranks reproducible SVIP RNA expression–survival associations across cancer types. High SVIP expression shows unfavorable associations in UCS and LIHC, but favorable associations in HNSC, KIRC, BRCA and OV. The HNSC Kaplan–Meier curve shows clear separation, with the low-expression group declining faster, consistent with the favorable association (log-rank p < 0.001). Together, the overview and detailed table identify HNSC as the clearest survival context for SVIP RNA expression.
This table summarizes SVIP tumor–normal expression differences by data type. RNA shows broader differences across cancer types, with a lineage consensus of 13, while mass-spec protein shows differences in 6. The strongest signals are observed in KIRC for RNA and CCRCC for protein.
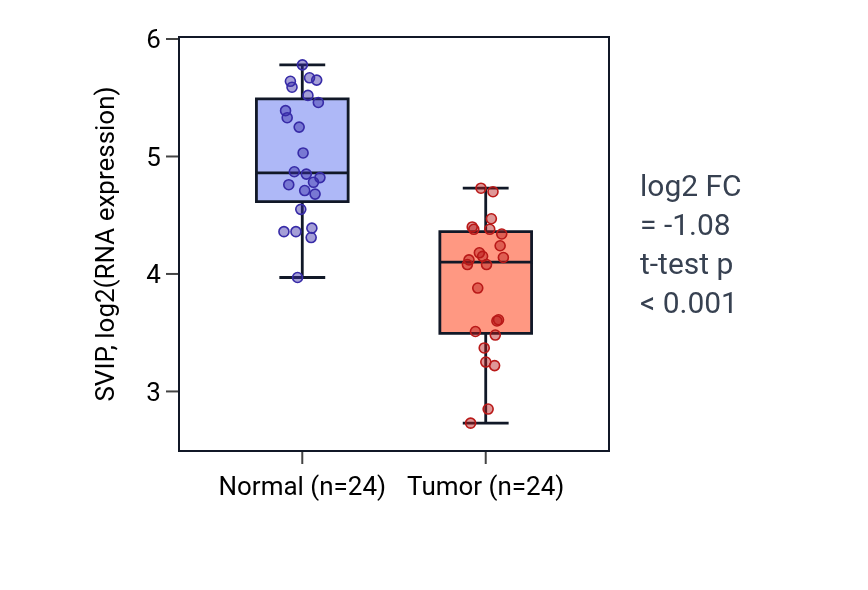
This table ranks reproducible tumor–normal expression differences for SVIP. A negative fold-change indicates higher expression in normal tissue than in tumor tissue. SVIP shows lower tumor expression in KIRC, HNSC, KIRP and LUSC and higher tumor expression in LIHC and BRCA. The KIRC box plot shows higher SVIP RNA expression in normal versus tumor tissue (log2 FC = −1.076, t-test p < 0.001).
This table shows molecular features associated with SVIP in patient tissues and cancer cell lines. In patient samples, SVIP shows the broadest associations at the RNA and protein expression levels, with GBM recurring as the lineage with the largest associated feature set. In cancer cell lines, SVIP RNA and mutation anchors are most strongly linked to RNA-expression features, especially in OVARY, while CRISPR and shRNA rows add functional-dependency signals in BREAST and BLOOD_Lymphoma.